Invited state-of-the-art review
The therapeutic potential of galanin in the management of pain – a review article
23. jan. 2024
18 min.
Abstract
Key Points
Karolinska Institutet has a long tradition in neuropeptide research. As early as 1931, Ulf von Euler discovered a molecule that influenced intestinal motility while working in the UK with John Gaddum, [1]. It was named substance P. In the Biochemical Department, Erik Jorpes and Viktor Mutt purified peptides from intestinal and brain tissues, among them cholecystokinin (CCK) [2]; and later vasoactive intestinal polypeptide (VIP) [3], neuropeptide Y (NPY) [4] and galanin [5]; and other peptides were discovered in the Mutt laboratory. These and many more peptides have been found to be expressed in neurons widely distributed in the central and peripheral nervous system. This work led to the idea of the existence of “peptidergic” neuron populations.
Peptidergic neurons were originally considered to be no different from other types of nerve cells, except for the messenger molecules. Thus, the peptidergic neurons simply seemed to represent a further neuron subpopulation, parallel to, e.g., monoaminergic and cholinergic neurons. However, a study on sympathetic ganglia revealed that a considerable population of neurons expressed somatostatin [6], i.e. the peptide originally discovered as a hypothalamic hormone [7]. Since virtually all neurons in the sympathetic ganglia are noradrenergic, the implication was that these neurons had somatostatin as a co-transmitter. Similar examples were found in the brain: many medullary serotonin neurons express substance P [8, 9] and most mesencephalic dopamine neurons synthesise CCK [10]. Moreover, it soon became clear that neuropeptides virtually always coexist with a classic transmitter and, conversely, neurons operating with monoamine or amino acid transmitters almost always express one or more peptides. This was a clarifying insight: it “degraded” neuropeptides to auxiliary messenger molecules and this, together with difficulties in establishing the functional role of peptides, e.g., in knock-out experiments, caused interest in neuropeptides to fade [11]. However, the question was asked: Is peptide the primary messenger of some neurons? In fact, some hypothalamic neurons synthesising the corticotropin-releasing hormone are GABAergic [12]; and many other hypothalamic systems, including the vasopressin and oxytocin neurons, are glutamatergic [13]. So here, peptides have a vital function and the amino acids are auxiliary messengers.
The multiple transmitter concept has grown increasingly complex; some neurons operate with several classic transmitters, e.g., dopamine with glutamate, adrenaline with glutamate, and glutamate with GABA [14]; and there are more combinations. Therefore, it became an even more complex task to explain how the co-released transmitters interact functionally. Also, it was clear that peptides are, in several ways, different from classic transmitters.
Neuropeptides are small proteins, i.e. their synthesis virtually always involves a gene and a transcript. Traditionally this process occurs in cell bodies and the peptide is then released from the nerve endings (Figure 1 A). This raised a number of important questions: Given the above characteristics, peptides would have to be transported over a distance of millimetres to meters (depending on system and species) to replace released peptide, and this could take up to days (Figure 1). This transport problem contrasts the situation for small-molecule transmitters, e.g., monoamines that may be synthesized in the nerve endings as synthesizing enzymes are also present there. Moreover, uptake sites (transporter molecules) exist at the membrane, allowing re-use of the transmitter. Possible explanations for this conundrum have emerged: synthesis and release can also occur in/from dendrites [15] (Figure 1 B) and, surprisingly, in/from nerve endings [16] (Figure 1 C). Importantly, peptides are slow transmitters (seconds to minutes and even much longer), are stored in a special type of vesicle, large dense core vesicles, and are released extrasynaptically following high/bursting activity [17, 18] (Figure 2 A and B). Peptides exert their actions at very low nanomolar concentrations via G protein-coupled receptors (GPCRs), and no reuptake mechanism is present at the cell membrane. Small-molecule transmitters are fast transmitters (milliseconds). They are stored in synaptic vesicles and are released already at normal firing rates (Figure 2 A and B).
Exactly 40 years have passed since the discovery of galanin [5], perhaps a good reason to highlight this peptide in this article. Animal research on galanin has generated many exciting options for relations to various diseases, suggesting a considerable translational potential. Here, we will focus on two aspects: involvement in pain and regeneration. The very broad panorama of functions of galanin and its underlying mechanisms were summarised by Lang et al. [19] and in 17 chapters in the book “Galanin” [20].
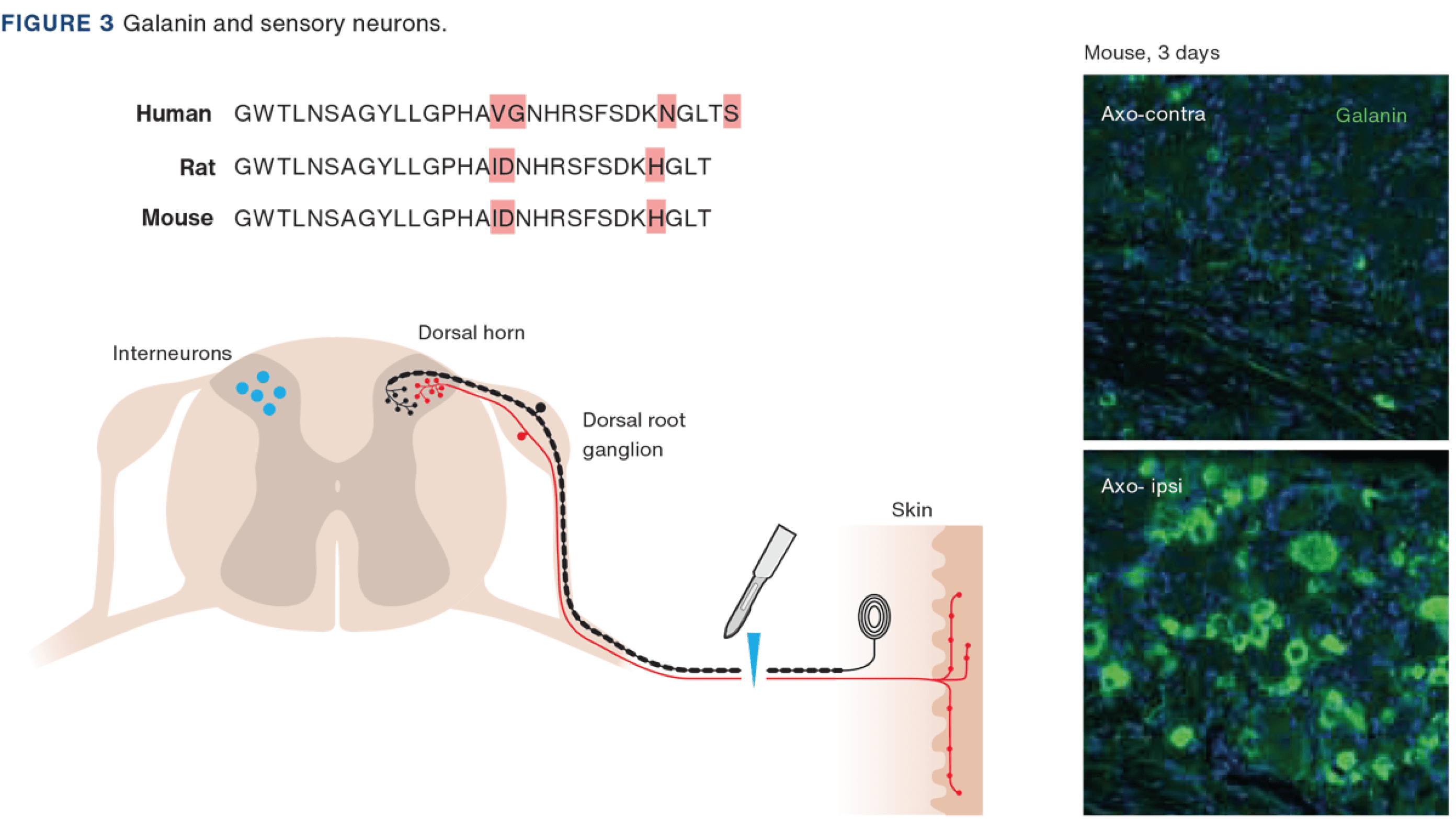
Galanin is a 29 (30 in humans) amino acid, C-terminally amidated peptide purified from porcine intestinal extract [5, 21]. The amino acid sequence is well preserved, including between human, rat and mouse (Figure 3), whereby the biological activity of rat galanin resides in the N-terminal portion; galanin(1-16) [22]. Today, four further peptides are considered to belong to the galanin family: galanin-message associated peptide [23], galanin-like peptide [24], alarin [25] and spexin [26]. Galanin exerts its action via three receptor subtypes: GalR1-3 [19].
In dorsal root ganglia (DRGs) of adult rats, galanin can be detected only at low levels under normal conditions [27, 28] (Figure 3). However, transection of the sciatic nerve causes a dramatic upregulation, up to 100 fold, of galanin synthesis [29, 30] (Figure 3). Interestingly, prenatally, galanin expression is very high in DRG neurons and is then downregulated and stays low during the rest of life [31], unless a “catastrophic” event occurs, like nerve injury. GalR1 and GalR2 are expressed in rat DRG neurons [32]. Of note, extensive electrophysiological, biochemical and histochemical evidence supports the claim that DRG neurons have glutamate as their principal transmitter [33]. Regarding galanin, at least two lines of research have evolved after these early findings, exploring a possible role of this peptide in i) pain signalling and ii) the survival and regeneration of sensory neurons.
Using the nociceptive flexor reflex models as read out, research has been conducted on the effect of galanin and galanin ligands on the nociceptive reflex in rats with or without peripheral nerve injury. These compounds had to be given intrathecally (ITH) as peptides only poorly penetrate the blood-brain-barrier. The results may be summarised as follows: In rats, galanin ITH exerts a biphasic effect on excitability with facilitation at low doses and depression at high doses [34-36]. The inhibitory effect is more pronounced after nerve injury [37, 38]. Subsequent studies indicated that the facilitation was mediated via GalR2 receptors, probably located in the DRG neurons, whereas GalR1 receptors, expressed in interneurons in the dorsal horn, were involved in the inhibition [39] (for details, see [40]).
Drawing on results from experiments on rats with peripheral nerve injury, we hypothesised that DRG neurons harbour an endogenous defence system combating pain, whereby the early pronociceptive effect was part of the beneficial “warning system”, whereas higher doses reduce neuropathic pain. This effect is enhanced by the parallel downregulation in DRG neurons of excitatory neuropeptides like substance P and calcitonin gene-related peptide (CGRP) [30]. This new idea complemented an already established mechanism attenuating inflammatory pain, involving dorsal horn interneurons expressing opioid peptides, among others [41]. These results obtained in rats also facilitated a suggestion for treatment: The early pain could be treated with a peripherally acting GalR2 antagonist and, importantly, the neuropathic pain by a GalR1 agonist acting in the spinal cord.
This said, it is important to note that peripheral nerve injury causes a dramatic effect on many more molecules in rodent DRG neurons: VIP [41] and NPY [42, 43] are also upregulated. Subsequently, microarray studies showed that hundreds of genes are regulated by nerve injury, and single-cell analyses of DRGs of several species were to follow. Thus, the galanin system is only one component of the DRG neurons that reacts to nerve injury, among others, aiming to defend against pain.
When discussing therapeutic aspects, the situation in humans needs to be considered. Recently, several single cell/nucleus studies have been published, but herein only the most recent one will be discussed [44]. Here, the galanin transcript is clearly detected in seven of the 16 neuron subpopulations, with the highest expression in transient receptor potential vanilloid-subtype 1 (TRPV1)/A1.1 and -/A1.2 groups, that is the peptide neurons expressing TRPV1 channels. However, neither GalR1, GalR2 nor GalR3 transcripts could be detected. Thus, our proposal that GaR2 is a target for treatment of the “initial” pain phase described above may not be relevant. Apparently the remaining, but also the most important, therapeutic target for neuropathic pain for galanin released from DRG neurons is therefore GalR1 receptors in the dorsal horn, requiring a GalR1 agonist.
Even if the upregulation of galanin after nerve injury obviously indicated a possible trophic role, it was a study by David Wynick and his group on a mouse with deleted galanin gene that provided the first evidence for such a function: this knock-out (KO) mouse had a 24% reduction in the number of substance P-positive neurons and a 15% reduction of the CGRP neurons [45], i.e. afflicting the small-diameter, nociceptive, C-fibre population. Also GalR2 KO mice exhibit a similar reduction of these neuron types [46, 47]. Moreover, nerve injury induces a marked loss of DRG neurons [48], and galanin KO and GalR2 KO mice exhibit diminished nociceptive behaviour [45, 49].
Galanin also promotes neurite outgrowth. Thus, following crush, injury regeneration is reduced by 36% in Gal KO mice compared with wild-type (WT) controls [45], and the compromised neurite outgrowth can be rescued with exogenous galanin [50]. For further detail, see [51]. Furthermore, the galanin KO mouse displays a 30% loss of cholinergic forebrain neurons [52], and galanin is upregulated in selective systems in the brain after injury [53]. These findings suggest that galanin may also have trophic effects in the brain. In fact, recently it was shown that overexpression of galanin in retinal ganglion cells promotes axonal regeneration after nerve crush [54].
Despite considerable efforts and several proposals being made in the course of four decades, no progress has been achieved in translating promising results from animal experiments and from analyses of human samples to medicines. A major obstacle has been the lack of success in developing small molecule, blood-brain-barrier-penetrating antagonists (or agonist), contrasting the situation for other peptides, such as members of the tachykinin family (substance P). In fact, only one such galanin antagonist has been generated, acting at the GalR3 receptor [55]. Moreover, as discussed above, the animal experiments may not provide proper guidance since human DRGs seem to lack both GalR1 and -R2.
The existence of more than a hundred neuropeptide GPCRs offers an, apparently, rich source of targets for drug development; not least because it is known that a large percentage of already available medicines acts via GPCRs. From that point of view, the outcome of some five decades of basic research on neuropeptides may appear meagre. However, some medicines were, indeed, approved by the US Federal Drug Agency (FDA), as summarised in Table 1: CGRP antibodies/antagonists are used to treat migraine [56]; orexin/hypocretin antagonists are sleeping pills [57]; NK1 (substance P) antagonists ameliorate chemotherapy-induced emesis [58]; GLP-1R agonists help diabetic patients, produce weight loss [59] and restore learning in obese individuals [60], and may perhaps even influence Alzheimer’s disease. Furthermore, NK3 antagonists reduce menopause-related flushes in women [61].
Based on Figure 2, showing the differential storage of peptides and the frequency dependence of release, the following hypothesis can be developed: Peptides are only released following high/bursting activity, which occurs during certain situations, e.g., a change in a physiological state, pain, stress or a disease [62]. The locus coeruleus (LC) neurons express many peptides (> 20), including galanin in ~ 80% of them [63]. It has now been recognised that in rodents, a compact LC nucleus encompasses several subgroups projecting to different brain regions with special functions [64]. Thus, selective activation of one subgroup may release galanin only in one specific brain region. Other galanin-containing LC neurons firing at low frequencies would release the main transmitter noradrenaline but not the peptide. A galanin antagonist would therefore block galanin signalling only in that activated region, without causing side effects related to the presence of galanin in nerve endings in other regions. Targeting peptide receptors may thus offer a high selectivity. A more extensive discussion of the galanin system in the LC and its relation to mood disorders was reviewed [65].
Neuropeptides represent a special class of neurotransmitters/messenger molecules, expressed in peripheral and central neurons. These systems are potential targets for drug development. In fact, several FDA approved drugs acting via a peptidergic mechanism are already on the market. Whether or not the galanin system will ever reach that status remains to be seen.
Correspondence Tomas Hökfelt. E-mail: tomas.hokfelt@ki.se
Accepted 10 November 2023
Conflicts of interest Potential conflicts of interest have been declared. Disclosure forms provided by the authors are available with the article at ugeskriftet.dk/dmj
Acknowledgements These studies were supported for six decades by the Swedish Research Council (last 2020-01688) and by the Wallenberg Foundations
Cite this as Dan Med J 2024;71(2):A10230653
doi 10.61409/A10230653
Open Access under Creative Commons License CC BY-NC-ND 4.0